
Génétique, Génomique, Epigénétique - ACTE II/II -"The Eighth Day of Creation", Horace Freeland Judson (1979) - "Biocode: The New Age of Genomics" 2015) - Siddhartha Mukherjee, "The Gene: An Intimate History" (2016) - "The Century of the Gene" (Evelyn Fox Keller, 2000) - Eva Jablonka & Marion J. Lamb, "Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life" (2005, 2014) - ...
Last update: 11/11/2025
De la génétique à l’épigénétique, en passant par la génomique, la science du vivant connaît moins une succession de ruptures qu’un déplacement progressif des formes d’intelligibilité du vivant.
La génétique installe un premier régime de savoir fondé sur le déterminisme héréditaire, où le vivant est pensé comme programme et l’individu comme héritage biologique.
La génomique complexifie ce schème en introduisant une pensée systémique et probabiliste : le vivant devient un réseau de corrélations, l’identité biologique un profil de risques calculables.
L’épigénétique, enfin, radicalise cette complexification en introduisant une plasticité régulée et historique, où l’expression du vivant dépend de l’environnement, des trajectoires individuelles et des conditions sociales.
Ces trois étages ne s’annulent pas : ils s’imbriquent.
Le déterminisme génétique n’est pas aboli, mais reconfiguré ; la complexité systémique n’est pas dépassée, mais historicisée ; la plasticité n’est pas liberté pure, mais vulnérabilité gouvernable. À mesure que le vivant devient plus plastique, il devient aussi plus étroitement gouverné.
Ainsi, la science du vivant ne transforme pas seulement la connaissance biologique : elle reconfigure la manière dont l’humain se pense lui-même.
L’individu n’est plus seulement déterminé, ni simplement calculé, mais engagé dans une gestion permanente de son avenir biologique. Le biopouvoir ne s’exerce plus seulement sur les corps, mais sur les conduites, les choix de vie et l’anticipation du risque.
La trajectoire Génétique–Génomique–Épigénétique révèle alors une tension centrale de la modernité biologique : plus le vivant est pensé comme modulable, plus il devient l’objet d’une normalisation diffuse et intériorisée ...
Le Gène, la Génétique ...
La génétique, discipline fondée sur les travaux de Gregor Mendel au XIXᵉ siècle, fondée sur l'ADN comme unité d’hérédité (publiés en 1866, redécouverts en 1900; 1910–1930, théorie chromosomique (Thomas Hunt Morgan) ; 1953, structure de l’ADN par James Watson et Francis Crick; à partir des années 1950, la génétique devient moléculaire), les gènes comme segments d’ADN codant des caractères : le gène est une unité identifiable, l’information génétique, vue comme séquentielle (suite de nucléotides), est stable, transmissible, codée.
Comment un caractère se transmet-il d’une génération à l’autre, quelle mutation est responsable d’une maladie, comment un gène influence-t-il un phénotype. Son niveau d'analyse est celui du gène individuel, son échelle la transmission, la mutation, la dominance ou la récessivité...

"The Eighth Day of Creation", Horace Freeland Judson (1979)
Ce n’est pas qu’un livre d’histoire des sciences, c’est le livre qui a inventé la manière dont on écrit l’histoire des sciences (des historiens comme Thomas Kuhn ou Robert K. Merton avaient certes déjà profondément transformé le champ). Paru en 1979, l’ouvrage d’Horace Freeland Judson s’est imposé comme un classique mondial de la révolution biologique du XXe siècle en donnant la parole à ses acteurs eux-mêmes. Il demeure, plus de quarante ans après sa publication, un incontournable absolu.
À la fin des années 1960, Judson n’était ni scientifique ni historien de formation, mais un journaliste culturel. Refusant de se contenter des publications officielles, il a entrepris une enquête de terrain sans précédent, 134 scientifiques interviewés, dont les plus grands (Crick, Monod, Jacob, Brenner, Perutz, Meselson…). Des entretiens répétés sur plusieurs années, avec confrontation croisée des témoignages. Un accès inédit aux correspondances privées et cahiers de laboratoire, permettant de vérifier la chronologie réelle des découvertes contre la mémoire parfois reconstruite des savants ..
Le livre couvre l’intégralité de la révolution moléculaire, des années 1930 aux années 1960 :
- La structure de l’ADN (1953), racontée non pas comme un coup de génie isolé, mais comme une pièce de théâtre avec ses dramatis personae.
- Le décryptage du code génétique et la découverte de l’ARN messager.
- L’invention de la cristallographie des protéines par Perutz et Kendrew.
Le génie de Judson est de rendre les scientifiques humains.
Loin des statues de bronze, ses personnages sont complexes, parfois admirables, parfois agaçants :
- Jacques Monod : Résistant, musicien, mais aussi « enfant gâté » qui préférait diriger une chorale à Pasadena plutôt que de travailler au labo.
- James Watson : Judson confronte la version du Double Hélix avec les lettres et notes de l’époque, révélant l’utilisation des données de Rosalind Franklin sans son consentement. Il réhabilite le travail de Franklin bien avant les biographies ultérieures (la réévaluation complète viendra plus tard).
- Sydney Brenner : Fils de cordonnier sud-africain, dont l’esprit vif et les lettres savoureuses traversent tout le livre.
Cette épaisseur psychologique transforme la science en drame humain.
En 2020, soit 40 ans après sa parution, le Molecular Biology of the Cell consacrait un éditorial à l’ouvrage. Son message : ce livre continue d’inspirer les vocations scientifiques. L’auteur de l’article raconte comment, étudiant indécis, la lecture de Judson l’a « mis sur une trajectoire de collision avec la science ». Réédité en 1996 puis en 2013 (édition commémorative), il reste l’introduction la plus vivante à la biologie moléculaire.
The Eighth Day of Creation de Horace Freeland Judson n’a pas été publié en français ...

Le Génome - La génomique, développée notamment avec le Human Genome Project (1990), développe une approche globale et systémique du génome en son entier (l'ensemble de l’ADN d’un organisme) : on découvre que les gènes n’agissent pas isolément, mais qu'il existe des réseaux, des interactions, des régulations croisées, et qu'une grande partie de l’ADN ne code pas directement des protéines. Combien de gènes possède un organisme, comment les gènes interagissent-ils entre eux, quelles variations génétiques existent entre individus, comment l’organisation globale du génome influence-t-elle la fonction, sont parmi les questions qu'elle se pose, son échelle est celle des interactions, des réseaux, des comparaison entre espèces ou individus.

"The Genome War : How Craig Venter Tried to Capture the Code of Life and Save the World", James Shreeve (2004) et "The Common Thread : A Story of Science, Politics, Ethics and the Human Genome", de John Sulston (2002) se situent tous deux dans le contexte plus large du débat sur la brevetabilité du vivant au début des années 2000.
James Shreeve est un journaliste et écrivain scientifique américain, spécialisé dans la vulgarisation des grandes découvertes en biologie, évolution et génétique. Il a écrit plusieurs ouvrages de science grand public, dont The Genome War et The Neandertal Enigma: Solving the Mystery of Modern Human Origin. Ses articles ont également été publiés dans des magazines reconnus comme The Atlantic Monthly, National Geographic, Science ou Smithsonian. "The Genome War" raconte la course intense entre deux approches pour séquencer le génome humain à la fin des années 1990 et au début des années 2000,
- Le projet public, soutenu par le gouvernement américain et mené par des institutions académiques (le Human Genome Project), qui visait à terminer le séquençage d’ici 2005.
- L’initiative privée de Craig Venter, avec sa société Celera Genomics, qui promettait d’achever le séquençage du génome humain bien plus tôt, en utilisant une méthode plus agressive (shotgun sequencing).
Shreeve y décrit la science, le business, les politiques, les intrigues et les personnalités impliquées dans cette compétition, mais il est le chroniqueur officiel du camp Venter et met en lumière des tensions réelles entre science publique et science privée, et montre comment l’annonce de Venter, visant à séquencer le génome humain rapidement (et potentiellement à en breveter des parties), a choqué une grande partie de la communauté scientifique traditionnelle.
Pour comprendre Celera, écrira un critique, lire "The Genome War", mais pour comprendre le projet public, autant lire "The Common Thread", de John Sulston...
John Sulston (1942-2018) était un biologiste britannique majeur du séquençage génomique, figure centrale du Human Genome Project. Prix Nobel de médecine en 2002 (pour ses travaux antérieurs sur le développement cellulaire de C. elegans), Il fut un défenseur très ferme du principe d’accès libre aux données génétiques (les fameuses « Bermuda Principles », publication immédiate des séquences). Son Récit autobiographique est engagé autour d'un argument central : le génome humain doit rester un bien commun mondial ...

"Biocode: The New Age of Genomics" (Dawn Field, 2015)
Ce n’est pas un manuel technique ou de référence scientifique mais un essai de synthèse prospectif sur ce que signifie entrer dans « l’ère du biocode » : l’idée que l’ADN de toutes les formes de vie devient une information lisible, stockable, partageable et exploitable à grande échelle. "Biocode" parce que toute la vie sur Terre peut être comprise comme un ensemble de codes numériques — des séquences d’ADN. Dawn Field nous explique ainsi cette révolution n’est pas seulement technique, mais qu'elle aussi épistémologique : elle transforme ce que signifie “connaître le vivant”...
Field propose un changement de perspective : le génome n’est plus seulement un objet biologique, mais devient une donnée globale, comparable à une infrastructure numérique. On est ainsi passé d'une biologie expérimentale traditionnelle à une biologie data-driven, fondée sur le séquençage massif et la bioinformatique.
Que se passe-t-il quand on peut séquencer tout ce qui vit — microbes, sols, océans, corps humain ?
- La biologie devient une science mondiale des données.
- La biodiversité invisible devient observable.
- La médecine change d’échelle.
- L’écologie devient quantifiable.
Mais, on ne peut omettre un fait d'importance : la compréhension reste plus lente que l’accumulation de données...
La biologie cesse d’être centrée sur quelques organismes modèles, elle devient une science de catalogage massif, l’ADN devient comparable à une base de données planétaire.
Le livre explique la chute spectaculaire du coût du séquençage, le passage du séquençage d’un génome unique à celui de millions d’organismes, et l’essor du séquençage environnemental (metagenomics). Celui-ci montre
- que l’immense majorité de la biodiversité est microbienne.
- que les écosystèmes sont gouvernés par des communautés génétiques invisibles.
- que les microbes structurent les cycles du carbone, de l’azote, du climat.
Conséquence : la biologie macro (plantes, animaux) n’est que la surface d’un monde génétique beaucoup plus vaste.
Si tout peut être séquencé, on change la manière de poser les questions scientifiques : on ne se demande plus seulement “Quel est cet organisme ?” mais “Quelles fonctions génétiques sont présentes ici ?”.L’unité d’analyse devient la fonction génétique, et non plus l’espèce.
Et si le séquençage ne donne pas immédiatement la compréhension fonctionnelle, le génome individuel, le microbiome, les variations populationnelles permettent, selon Field, une médecine personnalisée (prédictive?), une meilleure surveillance des maladies, une compréhension des interactions hôte–microbes.
Field suggère qu’avec le séquençage global, on peut surveiller la biodiversité en temps réel, on peut détecter les perturbations écologiques plus précocement, on peut cartographier l’évolution microbienne. Autrement dit, la planète devient “mesurable biologiquement”.
Reste que désormais, le vrai défi n’est pas tant de lire l’ADN, que de l’interpréter,et de l'interpréter collectivement. L'auteur insiste sur ces nouveaux défis que représente, par exemple, la surcharge massive de données ou les enjeux de gouvernance des données biologiques.
"... Nous restons submergés par l’immensité du Biocode de la Terre. Nous n’en avons encore vu qu’une infime partie. Miescher souhaitait à l’origine devenir prêtre, mais il entra finalement en école de médecine. Sa timidité et sa surdité rendaient impossible toute relation avec les patients, et il préféra le travail en laboratoire. C’est là qu’il découvrit la « nucléine ». Il fut surpris de la trouver non seulement dans le pus lavé de bandages, mais aussi chez le saumon. Beaucoup ont eu l’intuition que toute vie est connectée. Darwin a fourni la théorie la plus convaincante expliquant comment cela pouvait être le cas. Miescher fut le premier à voir physiquement ce qui rendait cette connexion possible : l’ADN. Grâce à Mendel, Watson, Crick et bien d’autres, nous connaissons aujourd’hui le mécanisme fondamental qui unifie la vie, confirmant la grande intuition de Darwin.
La génomique est à la vie ce que le tableau périodique est à la chimie. « Rien n’a de sens en biologie si ce n’est à la lumière de l’évolution » était le titre d’un article classique écrit par Theodosius Dobzhansky. En tant que généticien, Dobzhansky reconnaissait la contribution essentielle de l’ADN à la compréhension de l’évolution et à l’explication de la diversité et de l’unité du vivant. Mais la génomique ne peut pas répondre à toutes les questions que nous nous posons sur les organismes vivants ou les écosystèmes qu’ils forment. De même que la connaissance des éléments individuels ne suffit pas à expliquer toutes les propriétés des molécules complexes, la connaissance des génomes n’explique pas tout des organismes et des écosystèmes qu’ils constituent. De nombreux phénomènes sont « émergents » : ils apparaissent à des niveaux d’organisation plus élevés. Le moteur à combustion interne n’explique pas tous les comportements exaspérants de la circulation. Si les réponses aux questions des sciences de la vie ne s’arrêtent pas à l’ADN, elles commencent néanmoins par lui.
L’instance actuelle du Biocode de la Terre, le génome planétaire, nous inclut tous, ainsi que tous les autres organismes vivants. À partir du minuscule gland de LUCA, la première cellule, et de MILO, son premier génome, un immense arbre de la vie s’est développé jusqu’à dominer la planète. Sur des milliards d’années, il a formé les sols, rempli l’atmosphère d’oxygène et — par l’intermédiaire des humains — construit de grandes villes.
Il reste encore énormément d’ADN à explorer. La grande majorité des espèces et des gènes attendent d’être découverts. Ils codent des solutions aux défis auxquels les organismes vivants ont été confrontés au cours des éons de l’histoire de la Terre, y compris à des périodes où la planète connaissait des conditions radicalement différentes. Le génome planétaire représente un véritable trésor de composants qui stimulera l’industrialisation de la biologie synthétique. L’ingéniosité du vivant est contenue dans cet héritage génomique : c’est un capital génomique naturel.
La génomique ajoute des couches de détails d’une finesse remarquable à notre connaissance de la vie et de ses relations. La branche de l’arbre de la vie qui comprend les pigeons chers à Darwin, par exemple — un modèle de sélection artificielle — s’est récemment enrichie de façon spectaculaire. Darwin supposait que toutes les races domestiquées provenaient du pigeon biset. En 2013, les bases de données publiques de nucléotides sont passées de presque aucune séquence de pigeons à 38 génomes complets. Ces données ont montré de manière concluante que les pigeons bisets sont bien les ancêtres des plus de 1 200 races de pigeons d’ornement existantes — davantage que pour toute autre plante ou animal dans l’histoire humaine.
Le fait que tous les pigeons, et tous les organismes vivants, soient apparentés par descendance nous donne un avantage considérable pour comprendre la biologie de l’ensemble du vivant. Si cette unité est encourageante, l’immensité même de l’arbre inspire l’émerveillement. Nous savons qu’il existe des millions d’espèces qui n’ont pas encore été formellement décrites et d’autres encore à découvrir. Il y a plus de cellules microbiennes sur la planète que d’étoiles dans l’Univers. Elles représentent 50 % de la biomasse et 99 % de la diversité génétique sur Terre.
Nous ne sommes que des brindilles. Les analyses phylogénétiques, associées aux données archéologiques, anthropologiques et à d’autres types d’informations, montrent que les humains modernes sont apparus en Afrique il y a à peine 200 000 ans — l’une des espèces les plus récentes issues d’une procession de vie qui dure depuis plus de 3,5 milliards d’années. Nous nous trouvons à l’extrémité même d’un arbre qui ressemble davantage à une étoile en explosion, rempli de lignées unicellulaires de protistes aux ramifications profondes, ainsi que d’un ensemble tout aussi déroutant de bactéries et d’archées — pour la plupart encore non documentées. Certaines parties de l’arbre ressemblent en réalité davantage à un réseau, en raison du transfert « horizontal » de gènes entre des lignées parfois très éloignées, en particulier chez les microbes. Un jour, nous devrons peut-être ajouter un quatrième domaine : une vie extraterrestre provenant d’une autre planète, ou une vie synthétique que nous créerons ici sur Terre..." (Dawn Field and Neil Davies 2015)

Siddhartha Mukherjee, "The Gene: An Intimate History" (Scribner / Simon & Schuster, 2016)
Siddhartha Mukherjee est médecin oncologue et chercheur (Columbia University), essayiste de science à très large audience. Il est l'auteur à succès de "The Emperor of All Maladies", lauréat du prix Pulitzer.
Dans "The Gene", il nous propose une histoire intellectuelle et scientifique du concept de gène, depuis les spéculations antiques sur l’hérédité jusqu’aux développements contemporains de la génomique et de l’édition du génome. L’ouvrage ne vise pas à fournir un traité technique de génétique, mais à retracer la manière dont l’idée de gène s’est progressivement constituée, transformée et complexifiée au fil des avancées expérimentales et théoriques.
Le livre adopte une structure chronologique large, abordant successivement les conceptions pré-mendéliennes de l’hérédité, la génétique classique (Mendel, Morgan), la biologie moléculaire du XXᵉ siècle (structure de l’ADN, code génétique), puis les grandes entreprises contemporaines telles que le séquençage du génome humain et les technologies d’édition génétique. Mukherjee insiste sur le caractère instable et évolutif de la notion de gène, montrant comment celle-ci a été redéfinie à plusieurs reprises en fonction des outils expérimentaux disponibles et des cadres théoriques dominants.
Un trait distinctif de l’ouvrage est l’entrelacement du récit scientifique avec une réflexion personnelle ..
L’histoire familiale de l’auteur, marquée par des troubles psychiatriques, sert de point d’ancrage pour interroger les promesses et les limites de l’explication génétique des traits humains complexes. Cette dimension narrative n’a pas pour fonction de démontrer une thèse biologique nouvelle, mais de souligner les tensions persistantes entre déterminisme génétique, environnement et histoire individuelle, notamment lorsque les connaissances issues du laboratoire sont transposées dans les domaines médical, social ou politique.
Sur le plan conceptuel, "The Gene" reste globalement inscrit dans une vision gène-centrée modérée ..
Mukherjee reconnaît l’importance de la régulation, du contexte cellulaire et de l’environnement, mais ne développe pas une critique systématique du cadre de la Synthèse moderne ni une théorie alternative de l’hérédité (comme peuvent le faire, par exemple, Jablonka et Lamb). L’ouvrage se distingue davantage par sa capacité à mettre en perspective historique et humaine la génétique que par l’introduction de concepts théoriques nouveaux.
En ce sens, "The Gene" constitue moins un livre de débat théorique qu’un récit de vulgarisation savante, attentif aux enjeux éthiques, médicaux et sociaux de la génétique contemporaine, et particulièrement utile pour comprendre comment le gène est devenu une catégorie centrale — mais aussi problématique — de notre manière de penser l’hérédité et l’identité humaines.
"The Century of the Gene" (Evelyn Fox Keller, 2000)
Evelyn Fox Keller est considérée comme une figure majeure pour comprendre la transition du « siècle du gène » vers la biologie contemporaine. Physicienne de formation devenue historienne et philosophe des sciences, elle est surtout connue pour trois apports majeurs : son analyse critique du déterminisme génétique (elle s’oppose à l’idée simpliste selon laquelle un gène équivaut à un caractère, idée très répandue au XXe siècle, dans les médias comme dans une partie de la biologie), son travail sur les métaphores scientifiques (elle montre que la façon dont on parle des gènes (programme, plan, code, commande…) influence profondément la façon dont on pense la biologie — parfois au détriment de la réalité scientifique), et ses contributions aux études féministes des sciences(sans être militante au sens caricatural, elle analyse comment certains cadres conceptuels (contrôle, domination, hiérarchie) ont façonné la biologie moléculaire).
Evelyn Fox Keller propose ici une analyse historique et conceptuelle montrant que la notion de « gène » héritée du XXᵉ siècle est profondément ébranlée par les développements de la biologie moléculaire avancée et de la génomique. Son propos ne consiste pas à nier l’existence des gènes, mais à montrer que leur statut explicatif, leur délimitation et leur rôle causal ont été largement surestimés dans les cadres théoriques classiques.
1. Le gène n’est plus un objet discret
La génétique classique, issue de la synthèse mendélienne et de la biologie moléculaire des années 1950–1970, reposait sur une conception du gène comme unité bien définie, localisable et relativement autonome. Cette conception permettait d’établir des relations simples entre gènes, protéines et caractères phénotypiques.
Keller montre que cette vision est progressivement devenue intenable. Les recherches ultérieures ont révélé que ce que l’on appelle « un gène » peut produire plusieurs produits fonctionnels selon les conditions cellulaires (épissage alternatif), que des régions non codantes jouent un rôle déterminant dans l’expression, et que des segments d’ADN éloignés interagissent de manière coordonnée. Le gène apparaît alors moins comme un objet isolé que comme un ensemble de relations fonctionnelles, défini par un contexte expérimental et interprétatif.
2. Le gène n’a pas de frontières fixes
Un autre pilier de la critique de Keller concerne l’idée que les gènes posséderaient des limites naturelles clairement identifiables. En pratique, la génomique montre que les frontières entre gènes, régions régulatrices et séquences fonctionnelles sont souvent floues, variables et dépendantes du cadre d’analyse retenu.
Les promoteurs, enhancers et autres éléments régulateurs peuvent se situer très loin des régions codantes qu’ils influencent, parfois à l’intérieur ou au chevauchement d’autres gènes. Ainsi, la délimitation d’un gène n’est pas une donnée brute de la nature, mais une construction opératoire liée aux outils, aux hypothèses et aux objectifs des chercheurs.
3. L’action du gène dépend de multiples contextes
Keller insiste fortement sur le caractère contextuel de l’action génétique. Un même segment d’ADN n’a pas les mêmes effets selon le type cellulaire dans lequel il se trouve, le stade du développement, l’état physiologique de l’organisme, les conditions environnementales.
Le gène ne peut donc plus être conçu comme une cause linéaire et souveraine produisant directement un effet phénotypique. Il agit toujours au sein de réseaux complexes d’interactions moléculaires, cellulaires et environnementales. Cette dépendance au contexte invalide toute forme forte de déterminisme génétique simple.
4. La génomique et la « crise » de la notion classique de gène
L’un des paradoxes mis en évidence par Keller est que la génomique, loin de confirmer la centralité explicative du gène, en révèle les limites. Le séquençage massif des génomes a montré que le nombre de gènes est relativement réduit, que la complexité biologique provient largement de la régulation, de l’interaction et de l’organisation dynamique des systèmes biologiques, et non de la simple présence de gènes supplémentaires.

Ainsi, la génomique provoque une crise conceptuelle ....
non pas une crise de la biologie en tant que science, mais une crise des catégories héritées du XXᵉ siècle pour penser le vivant.
Le gène demeure indispensable, mais il ne peut plus être considéré comme l’unité fondamentale et suffisante de l’explication biologique.
Quand Keller parle de crise, elle ne parle ni d’échec scientifique, ni de remise en cause de l’ADN, ni d’un relativisme quelconque. Elle désigne une inadéquation croissante entre les catégories conceptuelles héritées du XXᵉ siècle et ce que les pratiques réelles de la biologie contemporaine révèlent. La science progresse, mais le langage explicatif hérité ne suit plus...
1. Les catégories héritées du XXᵉ siècle - La génétique classique et la biologie moléculaire ont cristallisé une image du vivant fondée sur quelques notions clés ...
a) Le gène comme unité fondamentale : unité de transmission, unité de fonction, unité de causalité
b) La causalité linéaire : ADN → ARN → protéine → caractère
c) Le programme génétique : le développement comme exécution d’un plan, l’organisme comme réalisation d’un code.
Ces catégories ont été extraordinairement fécondes. Elles ont permis la biologie moléculaire, le génie génétique, la médecine génomique précoce. Mais Keller montre qu’elles deviennent conceptuellement insuffisantes.
2. Ce que la génomique met concrètement en échec ...
2.1. L’unité du gène : la génomique révèle que ..
- un « gène » n’est pas une unité stable
- il n’y a pas correspondance 1:1 entre gène et fonction
- la fonction est distribuée entre régions codantes, régions régulatrices, structures chromatiniennes, interactions dynamiques
Si le gène reste indispensable, il ne porte plus à lui seul l’explication. C’est exactement ce que Keller appelle la fin du gène comme unité explicative suffisante.
2.2. L’effondrement de la causalité simple ..
Les systèmes génomiques montrent des boucles de rétroaction, des effets émergents, des causalités circulaires, des dépendances au contexte.
Un gène peut être ainsi causal dans un contexte, neutre dans un autre, ou compensé par un réseau ailleurs. On passe d’une causalité linéaire à une causalité systémique. C’est une transformation épistémologique, et non seulement technique.
2.3. Dès lors, le programme génétique devient une métaphore fragile ..
Keller insiste beaucoup sur ce point. La métaphore du programme suggèrait un contrôle central, une hiérarchie, une direction univoque. Or le développement réel est distribué, auto-organisé et dépendant de contraintes physiques, chimiques, environnementales.
C'est dire qie le génome participe au développement, mais ne le commande pas.
3. Mais ce n'est pas dire que nous sommes en pleine crise de la biologie ...
C’est crucial. La biologie contemporaine fonctionne progresse, produit des prédictions, transforme la médecine et l’écologie. La crise porte plutôt sur nos schémas explicatifs, nos métaphores, nos unités d’analyse héritées. Keller affirme (c'est l'idée centrale) que nous avons de meilleurs outils que jamais, mais des concepts devenus trop étroits.
Des ouvrages de références confirment ou prolongent Keller (The Century of the Gene) ...

- Keller, Evelyn Fox - "Making Sense of Life: Explaining Biological Development with Models, Metaphors, and Machines" (Harvard University Press, 2002)
Keller déplace la focale : au lieu de traiter le « gène » comme centre de commande, elle analyse comment on explique le développement (formation des formes, différenciation, stabilité des organismes) à l’aide de modèles (physiques, mathématiques), de métaphores (programme, information) et d’outils (machines expérimentales). Le livre insiste sur le fait que les explications biologiques robustes combinent souvent régulation génétique + dynamiques de systèmes + contraintes matérielles. C’est une continuation directe de The Century of the Gene, mais davantage orientée « épistémologie des explications » que « histoire du concept de gène ».

- Moss, Lenny - "What Genes Can’t Do" (The MIT Press, 2002 - hardcover; rééd. paperback 2004).
Moss reconstruit l’histoire du concept de gène et propose une distinction influente : le gène n’est pas (seulement) une « cause » au sens simple, mais plutôt une ressource mobilisée dans des processus développementaux et cellulaires. Il critique l’usage du gène comme explication « finale » (le gène fait le trait), et défend des explications où la causalité est distribuée (réseaux, contexte, développement). C’est très aligné avec l’idée kellerienne de crise : les catégories “gène = unité causale fondamentale” deviennent trop étroites.

- Oyama, Susan; Griffiths, Paul E.; Gray, Russell D. (eds.) - "Cycles of Contingency: Developmental Systems and Evolution" (A Bradford Book / The MIT Press, 2001).
Ce volume est une référence majeure de la Developmental Systems Theory (DST). L’idée : expliquer le vivant sans opposer “nature vs nurture” ou “gènes vs environnement”. Les traits biologiques se construisent via des cycles où interagissent ressources génétiques, environnementales, comportementales et sociales. Le livre propose donc une alternative au schéma gène→trait : le développement est un système distribué (et l’hérédité elle-même peut être pensée comme plurielle). Keller y figure d’ailleurs comme contributrice dans ce volume, ce qui illustre l’affinité intellectuelle.

- Neumann-Held, Eva M.; Rehmann-Sutter, Christoph (eds.) - "Genes in Development: Re-reading the Molecular Paradigm" (Duke University Press, January 2006).
Volume collectif explicitement conçu pour « relire » le paradigme moléculaire à la lumière de la génomique, du clonage, des cellules souches, etc. Il met en avant des phénomènes qui déstabilisent l’idée d’un gène comme unité explicative autonome (par ex. un même gène pouvant mener à plusieurs produits), et discute aussi les implications sociales/éthiques du déterminisme génétique. C’est une continuation très directe de la thèse de Keller : la génomique force à repenser le gène comme concept (et pas seulement comme objet technique).

"The Genome Factor : What the Social Genomics Revolution Reveals about Ourselves, Our History, and the Future", Dalton Conley , Jason Fletcher (2017, Princeton University Press)
Jason Fletcher est un économiste de la santé et un méthodologue quantitatif (Wisconsin). Dalton Conley est un sociologue public et théoricien (Princeton) : ensemble, ils ont contribué à fonder ce qu'ils appellent la "Social Genomics Revolution" (révolution de la génomique sociale) : l'idée que les sciences sociales ne peuvent plus ignorer les données génétiques, non pas pour retomber dans le déterminisme biologique, mais pour mieux mesurer et comprendre les effets de l'environnement.
L’objectif de Conley et Fletcher est bien de déclarer la fin de la guerre entre l'inné et l'acquis. Leur apport majeur ne se situe pas en biologie moléculaire, mais en épistémologie des sciences sociales.
- Le concept clé : Le "Molecular Me"
Le livre introduit l’idée que nous ne sommes plus obligés de nous contenter des études de jumeaux pour mesurer l’héritabilité. Grâce aux polygenic scores (scores polygéniques), les sociologues peuvent désormais intégrer des données génétiques réelles dans leurs modèles statistiques. Cela permet de ne plus considérer la génétique comme une "boîte noire".
- La déconstruction de l’hérédité (Chapitres 2 & 3)
L’ouvrage apporte une contribution technique importante en expliquant le "problème de l'héritabilité manquante" . Conley montre que si on ne trouve pas les gènes spécifiques de la réussite sociale, ce n’est pas parce qu’ils n’existent pas, mais parce que les effets sont polygéniques (des milliers de gènes avec des infimes effets) et interactifs (ils dépendent de l’environnement).
- Le "Tri génétique" et la mobilité sociale (Chapitres 4 & 6)
C’est peut-être la thèse la plus provocante du livre : le "cavorting" (le flirt/la cavalcade génétique). Conley démontre que nous ne choisissons pas nos conjoints uniquement sur des critères sociaux (diplôme, classe), mais aussi sur des critères génétiques (assortative mating). Ainsi, les inégalités se creusent car les "porteurs" de gènes favorables à l’éducation s’assemblent. Il applique cette logique à la macroéconomie en se demandant si une partie de la richesse des nations ne serait pas corrélée à la distribution de ces scores au niveau populationnel.
- La question raciale (Chapitre 5)
Contrairement aux dérives potentielles, le livre apporte une clarification importante : la race n’est pas génétique au sens biologique strict (pas de races humaines distinctes), mais la génomique permet de mesurer l’ascendance. Cette distinction permet de sortir du "nonsensical question" (la question absurde) de la race comme catégorie biologique, tout en étudiant scientifiquement les effets de l’ascendance et du racisme structurel.
The Genome Factor" est un livre de sociologie génomique, pas de sociologie épigénétique.
Il prend l'ADN fixe (le génome) et regarde comment il corrèle avec le comportement social. C'est une vision "statique" de la biologie et un livre sur la fin du déterminisme social pur. Il réintroduit le corps et l'ADN dans la sociologie.Cependant, il le fait sans sortir du logiciel néo-darwinien classique (l'ADN est le programme, l'environnement est le contexte).
L'épigénétique, au contraire, propose un modèle post-génomique où l'ADN n'est pas le chef d'orchestre mais une partition que l'environnement joue et réécrit...

"Blinded by science: The social implications of epigenetics and neuroscience" (David Wastell, Sue White, 2017)
C'est un livre de combat épistémologique et politique. David Wastell est professeur émérite de Systèmes d'Information à la Nottingham University Business School (Royaume-Uni); Sue White est professeure de Travail social à l'University of Sheffield (Royaume-Uni). Ils ne rejettent pas la biologie mais rejettent sa caricature. Leur contribution est unique, provocante, et profondément dérangeante pour le consensus scientifique dominant : "Pourquoi, se demandent-ils, cette science est-elle soudainement devenue si puissante dans les politiques sociales, alors que ses preuves sont si fragiles ?"
Part I: Getting to grips with the thought styles
- Biology and the drive for human improvement
L'idée d'"améliorer les humains par la biologie" n'est pas neuve. Un chapitre considéré comme le plus risqué pour la réputation du livre, parce qu'il établit un continuum entre l'eugénisme du XXe siècle et les politiques de "prévention précoce" du XXIe siècle. Mais il est vrai que sans cette généalogie, le lecteur ne comprend pas pourquoi il devrait s'inquiéter. Les promoteurs de l'Early Intervention ne se considèrent pas comme des eugénistes ; ils se considèrent comme des progressistes. Wastell/White doivent donc montrer que l'intention vertueuse n'empêche pas la dérive.
- How knowledge gets made in neuroscience and molecular biology
Il s'agit de montrer que la production de connaissances en neuroscience et en biologie moléculaire est incertaine, bricolée, dépendante de choix méthodologiques contestables. Les auteurs entendent démolir l'image d'Épinal de la "découverte" comme révélation soudaine. Wastell parle en praticien et peut expliquer pourquoi une expérience sur les rats ne se transpose pas mécaniquement aux humains; les effets statistiquement significatifs peuvent être biologiquement négligeables; les biais de publication (on ne publie que les résultats positifs) créent une illusion de robustesse; les corrélations entre méthylation et comportement sont presque impossibles à interpréter causalement. Ce chapitre est très technique. Wastell doit montrer les faiblesses sans tomber dans le scepticisme radical qui consisterait à dire que "rien n'est fiable". Or, sa position personnelle (il a quitté la neuroscience) le prédispose peut-être à ce biais.
- Blaming the brain
Les auteurs s'engagent sur le terrain de la critique du neuro-réductionnisme et entendent montrer comment des problèmes sociaux (pauvreté, échec scolaire, violence) sont re-définis comme des problèmes cérébraux. Leur analyse de la métaphore du "cerveau précaire" (precarious brain) porte sur l'idée que le cerveau de l'enfant pauvre est "câblé" différemment, de manière déficitaire. Habituellement, on présente les neurosciences comme un outil pour aider les enfants défavorisés ("comprendre leur cerveau pour mieux les éduquer"). Wastell/White retournent l'argument : en "biologisant" la pauvreté, on la naturalise. Ce n'est plus une injustice sociale à corriger ; c'est un déficit biologique à réparer. La réparation étant biologique, elle est confiée à des experts techniques (médecins, neuropsychologues), pas à des mouvements sociaux ou à des syndicats. La responsabilité de l'échec scolaire glisse de l'institution vers l'individu et sa mère (c'est elle qui n'a pas fourni un environnement utérin/stimulation précoce adéquats). Une analyse matérialiste puissante qui montre que le vocabulaire de la "bienveillance" masque un renforcement du contrôle social. Mais tous ceux qui s'intéressent au cerveau des enfants sont-ils des agents du néolibéralisme ? Et certains chercheur·ses en neurosciences éducatives sont-ils sincèrement animé·es par des intentions émancipatrices?
Pour se résumer, cette partie n'explique pas les mécanismes de l'épigénétique (méthylation, histones, etc.), ne propose pas d'alternative politique concrète, ne distingue pas entre les différents degrés de robustesse des recherches (tout est mis sur le même plan critique). Mais, et peu de livres le font, elle désacralise la biologie moléculaire, politise le débat sur l'épigénétique et tente d'armer intellectuellement les travailleurs sociaux, les parents, les citoyens face au discours de l'expertise neuroscientifique.
Part II: Fixing real people" - Un pamphlet brillant mais que de nombreux critiques estiment inabouti. Ainsi, par exemple,
- The precarious infant brain
Ce chapitre opère une archéologie du concept de "cerveau précaire". Wastell/White montrent que l'idée selon laquelle le cerveau infantile est irréversiblement vulnérable aux expériences précoces n'est pas une découverte neuroscientifique récente, mais la résurgence d'un mythe des années 1970 (la "théorie des dommages cérébraux sociogéniques" de Valentine et al., 1975). La nouveauté n'est pas la thèse, c'est son emballage technologique : l'imagerie IRMf remplace les hypothèses spéculatives d'antan et renforcer le rapport Allen (2011), texte fondateur des politiques d'Early Intervention au Royaume-Uni (un texte politique, mais non scientifique).
- The cat is out of the bag: from early intervention to child protection
Un chapitre tenu par nombre de critiques comme le plus dérangeant politiquement. Wastell/White démontrent que le même discours neuroscientifique qui justifie l'intervention précoce bienveillante justifie aussi les mesures coercitives de retrait d'enfant ("forced adoption"). Il n'y a pas deux politiques opposées (gauche/droite, soutien/répression) ; il y a un seul continuum narratif : la science du cerveau vulnérable légitime toute intrusion étatique, quel que soit son degré de coercition. ils exhument le discours de David Cameron (11 janvier 2016) où le Premier ministre conservateur utilise explicitement les travaux de Jack Shonkoff (Harvard) pour justifier à la fois les visites à domicile ET les adoptions sans consentement. Ce n'est pas un argument complotiste, les faits rapportés sont disponibles; ils montrent comment la métaphore du "tennis" ("mums and dads build babies' brains") réduit la parentalité à une performance technique évaluable, ouvrant la voie à la sanction en cas de "mauvais jeu". Mais nos auteurs ne proposent pas de critère pour distinguer une "bonne" traduction de la science d'une "mauvaise". Toute mobilisation politique de la neuroscience devient suspecte par contamination.
- Epigenetics: rat mum to my Mum?
C'est le cœur technique du livre sur l'épigénétique. Wastell/White attaquent frontalement les travaux de Michael Meaney et collègues (l'exemple canonique de l'épigénétique comportementale : les rates qui lèchent leurs petits produisent une progéniture moins stressée, via méthylation du récepteur aux glucocorticoïdes). Deux niveaux de critique sont élaborées, critique interne (failles méthodologiques : tailles d'échantillon insuffisantes, effets statistiquement significatifs mais biologiquement négligeables) et externe (transposition interspèce : même si le modèle fonctionnait chez le rat, sa généralisation à l'humain est épistémologiquement injustifiée; la complexité des pratiques parentales humaines n'a aucun équivalent dans le léchage/épouillage). C'est démonter l'expérience la plus médiatisée de l'épigénétique. Mais si l'argumentaire Meaney tombe, toute l'épigénétique comportementale humaine s'effondre-t-elle ? Le chapitre ne le dit pas. Depuis, certains aspects des travaux de Meaney ont été partiellement répliqués, d'autres non. Le chapitre est un instantané, pas une méta-analyse définitive.
- Human epigenetics prematurely born(e)?
Ce chapitre opère le basculement du rat à l'humain. Il examine la littérature sur l'épigénome humain, en particulier les études sur :
- les catastrophes naturelles (famine hollandaise, 1944-45) – le cas emblématique de l'épigénétique environnementale.
- le paradigme DOHaD (Developmental Origins of Health and Disease).
L'épigénétique est ici présenté en fin de compte comme un instrument de gouvernement des populations, et non comme un corpus de connaissances neutres ..
Le livre identifie un coupable spécifique, ignoré par tous les autres ouvrages : le modèle DOoHD (Developmental Origins of Health and Disease) selon lequel l'environnement précoce (notamment in utero et petite enfance) programme de manière durable et souvent irréversible le cerveau et le métabolisme. Le modèle a émergé progressivement des travaux de David Barker (1938-2013), épidémiologiste britannique à l'Université de Southampton, pour être systématisé en 1992 et être institutionnalisé en 2000-2001. Les années 2005-2010 virent l'ntégration des mécanismes épigénétiques comme plausibilité biologique (méthylation, modifications d'histones, ARN non codants), les pathologies concernées furent étendues (au cardiovasculaire et diabète furent aloutés obésité, cancer, maladies rénales chroniques, ostéoporose, troubles neuropsychiatriques, asthme, fertilité), et les expositions élargies, de la malnutrition au stress maternel, tabagisme, pollution atmosphérique, perturbateurs endocriniens, diabète gestationnel, obésité maternelle.Il faut observer que le DOHaD est devenu un langage commun international (du Royaume-Uni aux Etats-Unis, Canada, Inde, Japon, Chine, Afrique du Sud). Et que le DOHaD contemporain utilise massivement des catastrophes naturelles et humaines comme "expérimentations naturelles" hors d'Europe (génocide rwandais, famine chinoise, etc, cf.Journal of Developmental Origins of Health and Disease).
Wastell/White montrent que ce modèle, présenté comme une découverte neuroscientifique révolutionnaire, est en fait une doctrine politique déguisée en science. Il légitime : la surveillance des femmes enceintes ("l'environnement utérin"), les interventions précoces obligatoires, le transfert de responsabilité de la société vers la mère.
Le DOHaD n'est PAS une hypothèse britannique des années 1980, ce n'est pas non plus une théorie fixe, mais une boîte à outils rhétorique dont la plasticité conceptuelle permet des usages politiques très divers, y compris ceux qu'ils critiquent : et c'est, depuis 2001, le langage scientifique PLANÉTAIRE pour penser les origines développementales des maladies et la santé publique internationale contemporaine. C'est précisément pour cela que sa critique par Wastell/White est politiquement explosive....
Mais si le DOHaD est une idéologie, comment penser les véritables effets de la malnutrition fœtale ? Le sous-titre du livre annonçait "social implications", mais les implications pratiques pour les travailleurs sociaux, les décideurs ou les citoyens restent floues ...
Au moins nos auteurs apportent-ils une mise en garde ...
Rick Hood, dans sa recension pour le British Journal of Social Work, pose la question que Wastell/White osent formuler : "Are we sleepwalking into a new era of state-sanctioned epi-eugenics?" L'eugénisme historique justifiait l'élimination des "inaptes" par la génétique (hérédité fixe). L'"épi-eugénisme" justifie l'intervention précoce et le façonnement des populations par l'épigénétique (hérédité modifiable). Le vocabulaire change, la logique de contrôle reste? Le livre établit cependant que les conditions discursives (parler de "cerveau précaire", de "programmation fœtale", de "transmission intergénérationnelle de la pauvreté par méthylation") sont déjà en place ..

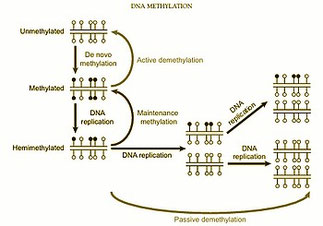
Epigénétique, la Régulation des gènes ...
L’épigénétique, concept formalisé par Conrad Waddington au XXᵉ siècle (1942), a pour socle l’ADN comme support stable mais son niveau d’analyse est celui de la régulation de l’expression des gènes. L’ADN ne change pas, mais son accessibilité et son expression varient, son expression est modulée par des marques chimiques réversibles et contextuelles, l’environnement entre dans le jeu. L’information génétique devient contextuelle, dynamique, historique.
- 1942 - Naissance du mot : Conrad Hal Waddington (biologiste britannique) crée le terme "épigénétique". Il veut fusionner "épigenèse" (développement progressif de l'embryon) et "génétique". Il imagine un "paysage épigénétique" pour expliquer comment un organisme se développe à partir d'un œuf .
- 1958-1962 - La cristallisation théorique (La "vraie" naissance) : Ce n'est pas Waddington qui fonde l'épigénétique moderne. C'est le dogme de la biologie moléculaire lui-même qui l'a rendue nécessaire. On découvre que toutes les cellules contiennent le même ADN. Dès lors, des mécanismes inconnus doivent exister pour activer ou éteindre les gènes. Le concept est donc déduit avant d'être observé .
- François Jacob & Jacques Monod - Modèle de la régulation génétique (opéron lactose). Pose la question de "l'induction" environnementale (1962)
- John Gurdon - Transplantation nucléaire : preuve que le noyau d'une cellule différenciée contient tout le génome (1962)
- 1975 - L'hypothèse moléculaire : Robin Holliday et Arthur Riggs postulent que la méthylation de l'ADN est le mécanisme responsable de cette transmission de l'activité des gènes .
- 1999-2003 - Les preuves expérimentales (Ancrage définitif) ..
- 1999 : Emma Whitelaw montre qu'une modification épigénétique (couleur du pelage) peut passer à la descendance chez la souris .
- 2003 : Randy Jirtle et Robert Waterland (Université Duke) réalisent l'expérience de la souris "Agouti". En nourrissant des souris gestantes avec des donneurs de méthyle, ils modifient le phénotype (couleur, santé) de la descendance sans changer l'ADN. C'est l'expérience la plus citée comme "preuve de concept" de l'influence environnementale
Pourquoi deux cellules avec le même ADN (ex. neurone et cellule musculaire) se comportent-elles différemment ? Comment l’environnement (stress, alimentation, toxiques) influence-t-il l’expression des gènes ? Certaines modifications acquises peuvent-elles être transmises ? Telles sont les quelques interrogations qu'elles se posent à l'échelle de la méthylation de l’ADN, de la modifications des histones, de la structure de la chromatine.
Dans les années 2000, l’épigénétique devient centrale parce qu’elle ....
- explique comment un même génome produit des cellules, tissus et comportements différents
- éclaire des pathologies complexes sans mutation génétique
- fournit des cibles thérapeutiques réversibles
- et transforme notre conception du génome, de statique à hautement régulé et contextuel

L’épigénétique s'impose dès lors en biologie du développement, cancérologie, et neurosciences, avec ...
1. - La découverte du rôle central de la méthylation de l’ADN
La méthylation de l’ADN correspond à l’ajout de groupements méthyle (–CH₃), le plus souvent sur les cytosines des dinucléotides CpG. Dans les années 1990–2000, on comprend que la méthylation réprime l’expression des gènes (gènes « éteints »), qu'elle est héritable lors des divisions cellulaires, sans modifier la séquence d’ADN. Et qu'elle joue un rôle clé dans la différenciation cellulaire (même ADN, cellules différentes), l’empreinte parentale, l’inactivation du chromosome X, la cancérogenèse ("silenciation" de gènes suppresseurs de tumeurs). En cancérologie, on découvre que des profils de méthylation anormaux sont aussi importants que les mutations génétiques.
2. - Les modifications des histones
L’ADN est enroulé autour de protéines appelées histones, formant la chromatine. Dans les années 2000, on met en évidence un véritable « code des histones » ..
- acétylation → chromatine ouverte → gènes actifs
- méthylation → activation ou répression selon le site
- phosphorylation, ubiquitination, etc.
Ces modifications modulent l’accessibilité de l’ADN, sont dynamiques et réversibles et permettent une régulation fine, rapide et spécifique de l’expression génique
En biologie du développement, cela explique comment des programmes génétiques complexes sont activés ou réprimés dans le temps et l’espace.
3. - ARN non codants ...
La vision « un gène = une protéine » s’effondre au début des années 2000. On découvre de nombreux ARN non codants : microARN (miRNA), siRNA, lncRNA.
Leur rôle est de définir la régulation post-transcriptionnelle, le guidage de complexes épigénétiques, la silenciation ciblée de gènes, l'organisation de la chromatine.
En neurosciences, ces ARN sont cruciaux pour la plasticité synaptique, la mémoire et le développement cérébral.
4. - Les projets comme ENCODE
Le projet ENCODE (Encyclopedia of DNA Elements), lancé au début des années 2000, marque un tournant majeur. Ses résultats clés ...
- une grande partie du génome est fonctionnelle, même hors des gènes codants
- une cartographie massive des régions régulatrices, des marques épigénétiques, des interactions ADN-protéines
- l'intégration des données épigénétiques à grande échelle ENCODE institutionnalise l’épigénétique comme pilier de la biologie moderne, au même titre que la génomique.

"Epigenetics"(2007, C. David Allis et al.)
C'est le premier grand manuel académique de référence en épigénétique. A sa parution en 2007, ce livre est exceptionnel parce qu'il il synthétise un champ jusque-là éclaté (méthylation de l’ADN, chromatine, ARN non codants, développement, cancer), et qu'il est dirigé par des figures fondatrices du domaine, C. David Allis (code des histones), Thomas Jenuwein et Danny Reinberg. il établit une définition opérationnelle moderne de l’épigénétique et sert rapidement de manuel de base en master/doctorat. Dans les années 2007–2012, c’est le livre que l’on cite quand on veut montrer que l’épigénétique est devenue une discipline structurée.
Mais il précède l’explosion du séquençage haut débit, la maturation des données ENCODE, l’essor massif des lncRNA, l’épigénétique mono-cellulaire et reflète une vision encore très chromatine-centrée. Certaines notions ont été depuis redéfinies ou nuancées. On associe souvent Allis (2007) à des éditions ultérieures (2015, 2023), des revues de synthèse post-ENCODE et des manuels intégrant l’épigénétique des systèmes complexes.

"Epigenetics Revolution" (Nessa Carey, 2011)
"Epigenetics: How Environment Shapes Our Genes", de Richard C. Francis, est le premier livre destiné au grand public sur le sujet de l'épigénétique. Narratif et anecdotique, Francis utilise des histoires fortes (la famine hollandaise, les stéroïdes et José Canseco, les diables de Tasmanie) pour "planter des drapeaux" conceptuels. La citation de Time ("The age of epigenetics has arrived") mise en avant par l'éditeur n'est pas un argument marketing creux : elle ancre ce livre comme le symptôme et le vecteur de l'émergence du sujet dans la conscience publique. Il sera encore lu et commenté en 2023-2024 car Francis est un biologiste du développement. Il s'intéresse à "ce que ça signifie" pour la théorie de l'évolution.
Mais c'est "Epigenetics Revolution" qui va s'imposer comme l’un des premiers livres de vulgarisation scientifique à avoir rendu accessible au grand public la complexité de l’épigénétique : les métaphores de Carey sont souvent saluées comme les meilleures du genre et elle ne parle jamais de concepts sans décrire l’expérience qui a permis de les découvrir. Si son approche est souvent saluée pour sa clarté analogique, mais critiquée pour certaines simplifications.
Écrit en 2011, le livre, c'est la difficulté de l'exercice, ne traite pas les révolutions récentes (CRISPR pour l’épigénome, CRISPRoff, cartographie unicellulaire de l’épigénome) et sous-représnte les ARN non codants : à l’époque, on sous-estimait encore le rôle central des longs ARN non codants (lncRNA) dans le recrutement des complexes épigénétiques. Reste que la philosophie du livre (l’interaction entre l’environnement et nos gènes est continue et dynamique) est plus pertinente que jamais.
Chapitre 1 : Un crapaud laid et un homme élégant
Ce chapitre d’ouverture plante le décor en opposant deux figures majeures de la biologie du développement.
Le crapaud laid : Il s’agit des expériences de John Gurdon en 1958. Il a pris le noyau d’une cellule intestinale de têtard, l’a inséré dans un ovule énucléé, et a obtenu un clone (un crapaud). Cela prouve que le noyau d’une cellule différenciée contient toujours le génome complet et intact. La différenciation cellulaire n’est donc pas une perte de gènes.
L’homme élégant : Il s’agit de Conrad Waddington, père du terme "épigénétique". Il a imaginé le paysage épigénétique : une métaphore où une cellule est une bille qui descend une pente. Au début, toutes les billes sont au sommet (cellules souches), puis elles empruntent des vallées (voies de différenciation) qui deviennent de plus en plus profondes, rendant le retour en arrière impossible.
Carey utilise ici une narration romancée très efficace. Cependant, certains critiques notent qu’elle simplifie le débat historique. À l’époque de Gurdon, beaucoup pensaient encore que le clonage était impossible chez les mammifères (Dolly n’arrivera qu’en 1996). Le chapitre pose parfaitement la question centrale du livre : Si toutes nos cellules ont le même ADN, pourquoi sont-elles différentes ?
Chapitre 2 : Comment nous avons appris à remonter la pente
Carey explique ici le mécanisme qui bloque les cellules dans leur vallée.
Elle introduit la méthylation de l’ADN. C’est une modification chimique (ajout d’un groupement méthyle sur une cytosine) qui agit comme un "verrou" sur le gène. Plus un gène est méthylé dans sa région promotrice, moins il est exprimé.
Elle utilise l’exemple des cellules B et T du système immunitaire. Ces cellules possèdent le gène pour fabriquer des anticorps, mais seules les cellules B le font. Dans les cellules T, ce gène est méthylé et donc éteint de manière stable.
La "remontée de pente" : les chercheurs ont réussi, via des drogues déméthylantes, à "déverrouiller" des gènes et à reprogrammer des cellules.
Le chapitre est très didactique. La limite ici est que Carey ancre presque exclusivement l’épigénétique dans la méthylation. En 2011, on savait déjà que d’autres mécanismes (modifications des histones, ARN non codants) étaient aussi centraux. Cette focalisation est compréhensible pour la clarté, mais peut donner au lecteur une vision trop "binaire" (méthylé = éteint, déméthylé = allumé), alors que la réalité est bien plus nuancée.
Chapitre 3 : La vie telle que nous la connaissions
Ce chapitre fait un rappel des fondamentaux de la génétique "classique".
Carey revient sur la structure de l’ADN, le dogme central (ADN → ARN → Protéine), et le code génétique. Elle insiste sur la différence entre génotype (la séquence) et phénotype (l’expression visible). L’objectif est de montrer que la génétique seule ne peut pas expliquer la complexité des organismes (notamment le nombre de gènes humains, seulement ~20 000, autant qu’un ver).
Ce chapitre est un passage obligé. Il est bien construit mais peu innovant. Il sert de "Socle" pour le lectorat non-scientifique. La critique que l’on pourrait faire est qu’il entretient parfois l’idée d’un "tout génétique" contre lequel l’épigénétique viendrait se battre, alors que les deux champs sont profondément intriqués.
Chapitre 4 : La vie telle que nous la connaissons maintenant
C’est la révolution épigénétique proprement dite.
Carey introduit le concept de chromatine. L’ADN n’est pas nu ; il est enroulé autour d’histones (formant des nucléosomes). Elle explique les modifications des histones (acétylation, phosphorylation). Si la méthylation de l’ADN est un verrou, les queues d’histones sont des "interrupteurs" ou des "panneaux indicateurs". Elle utilise la métaphore de la bibliothèque : L’ADN est le livre, les histones sont le bibliothécaire qui décide de mettre le livre en accès libre, en réserve ou à la poubelle.
C’est le cœur technique du livre. Carey est brillante dans ses métaphores. Cependant, elle simplifie le code histone. En 2011, on découvrait que ces modifications sont extrêmement complexes (certaines activent, d’autres répriment selon le contexte). Le livre donne l’impression que le système est plus cartésien qu’il ne l’est vraiment.
Chapitre 5 : Pourquoi les vrais jumeaux ne sont-ils pas vraiment identiques ?
C’est l’un des chapitres les plus célèbres du livre. Les jumeaux monozygotes partagent le même ADN. Pourtant, l’un peut développer une schizophrénie, l’autre non ; l’un peut être obèse, l’autre mince. Carey explique que cela est dû à une dérive épigénétique. Au fil du temps, l’exposition à des facteurs environnementaux (alimentation, tabac, stress) ajoute ou enlève des marques épigénétiques. À 3 ans, les jumeaux ont des épigénomes quasi identiques. À 50 ans, ils sont radicalement différents.
Ce chapitre est extrêmement puissant pour le grand public car il répond à une question intuitive. La critique scientifique récente (post-2011) nuance toutefois cet effet. Des études plus larges montrent que l’environnement in utero est souvent plus déterminant que l’environnement postnatal précoce. De plus, une part importante de la variation épigénétique reste aléatoire (bruit de fond), ce que Carey sous-estime.
Chapitre 6 : Les péchés des pères
Carey aborde la transmission transgénérationnelle.
Étude clé : Överkalix, en Suède. Les historiens ont étudié les récoltes des grands-pères et la mortalité cardiovasculaire des petits-enfants.
Résultat frappant : Si un grand-père a vécu un pic de famine (mauvaise récolte) juste avant sa puberté, ses petits-enfants vivent plus longtemps et meurent moins de maladies cardiaques. Si le grand-père a trop mangé (bonnes récoltes), ses petits-enfants meurent plus jeunes du diabète.
Le "péché" (l’excès de nourriture) est "transmis" via les spermatozoïdes sous forme de marques épigénétiques.
Chapitre fascinant mais controversé. Carey le présente avec beaucoup de certitude. La communauté scientifique est plus prudente : il s’agit d’une étude rétrospective avec un échantillon statistique relativement faible. De plus, le mécanisme exact (comment une marque épigénétique survit-elle à la reprogrammation massive qui a lieu dans l’embryon ?) reste mal compris. Carey omet de dire que ce type de transmission est rare et bien moins robuste que chez les plantes ou le ver.
Chapitre 7 : Le jeu des générations
Ce chapitre explore la reprogrammation épigénétique.
Chez les mammifères, juste après la fécondation, TOUTES les marques épigénétiques sont effacées (sauf quelques exceptions). Puis, un peu plus tard, elles sont "réécrites" selon le sexe de l’embryon et le type cellulaire. Carey utilise l’exemple des souris Agouti. Ces souris sont génétiquement identiques mais de couleur différente (jaune obèse ou brune mince) selon le régime alimentaire de leur mère (supplémentation en donneurs de méthyle).
Ce chapitre est un chef-d’œuvre pédagogique. Il prouve que l’épigénétique n’est PAS un "lamarckisme" pur et simple, puisqu’il y a un effacement systématique. L’hérédité épigénétique est une "fuite" dans ce système de nettoyage, pas la règle. C’est une nuance cruciale que Carey expose très bien.
Chapitre 8 : La bataille des sexes
Ici, Carey traite de l’empreinte génomique (genomic imprinting). Certains gènes ne s’expriment que s’ils viennent du père, d’autres que s’ils viennent de la mère.
Exemple : IGF2. Si la copie maternelle est active et la paternelle éteinte, l’embryon est trop petit. Si l’inverse se produit, l’embryon est trop gros.
Théorie évolutive : Le père veut des bébés gros pour assurer sa descendance (quitte à épuiser la mère), la mère veut des bébés petits pour survivre à l’accouchement et préserver ses ressources pour d’autres grossesses.
Carey excelle dans la vulgarisation de concepts évolutifs. Cependant, la vision du "conflit parental" est très anthropomorphique. De plus, elle néglige le fait que l’empreinte est un mécanisme risqué (une seule mutation sur le gène actif cause la maladie) et que beaucoup de ces mécanismes semblent être des "accidents" évolutifs plutôt que des stratégies parfaitement rodées.
Chapitre 9 : Génération X
Ce chapitre parle du chromosome X et de son inactivation. Les femmes (XX) n’ont pas deux fois plus de protéines des gènes du X que les hommes (XY). Pourquoi ? Car un des deux X est totalement silencieux. Ce silence est orchestré par un ARN non codant appelé Xist. Xist "recouvre" un chromosome X comme une couverture, et recrute des enzymes pour le compacter en hétérochromatine (corps de Barr). C’est aléatoire : dans une cellule, c’est le X du père qui est éteint, dans la cellule d’à côté, celui de la mère. D’où le motif "tacheté" des chats calico.
Chapitre très solide. Carey montre ici l’importance des ARN non codants, qu’elle avait peu abordés jusqu’ici. C’est un contrepoint intéressant à son obsession pour la méthylation.
Chapitre 10 : Le message n’est pas le médium
Carey aborde les maladies complexes et la difficulté de trouver des "gènes responsables". La plupart des maladies ne suivent pas les lois de Mendel (un gène = une maladie). Elle explique que les études d’association pangénomique (GWAS) échouent souvent à trouver des gènes responsables pour les maladies psychiatriques ou auto-immunes.
L’épigénétique est proposée comme le "chaînon manquant" entre la prédisposition génétique et le déclenchement environnemental.
C’est une critique assez juste de la génétique des années 2000. Cependant, Carey tombe parfois dans le piège inverse : faire de l’épigénétique la réponse à tout. La réalité est que beaucoup de maladies complexes sont polygéniques (des centaines de gènes avec de très petits effets) plutôt que principalement épigénétiques.
Chapitre 11 : Combattre l’ennemi intérieur
Ce chapitre est consacré aux rétrovirus endogènes. 8% de notre génome est constitué de virus anciens qui se sont insérés dans notre ADN il y a des millions d’années. Si ces virus s’exprimaient, ils seraient toxiques ou infectieux. Heureusement, ils sont maintenus sous silence par la méthylation de l’ADN.
Carey explique que les cellules souches embryonnaires ont un système de répression encore plus strict (complexe Polycomb) pour garder ces virus silencieux tout en gardant les gènes de développement "prêts à l’emploi".
Chapitre brillant et original. Il montre que l’épigénétique est un système de défense antivirale évolué. On touche ici à la notion de "génomique conflictuelle".
Chapitre 12 : Tout dans la tête
Application de l’épigénétique aux neurosciences.
Comment le cerveau stocke-t-il des mémoires pendant des décennies alors que les protéines et les ARN ne vivent que quelques heures ? Réponse : l’épigénétique. L’apprentissage provoque des modifications des histones et de la méthylation qui stabilisent les connexions neuronales.
Elle prend l’exemple du Rett syndrome, une maladie liée à une mutation du gène MeCP2 (une protéine qui lie l’ADN méthylé). Sans MeCP2, le cerveau ne peut pas maintenir ses synapses.
Carey est très prudente ici, car en 2011, le champ de l’épigénétique comportementale était naissant (et parfois controversé, notamment les travaux de Michael Meaney sur le léchage des rats). Elle évite le sensationnalisme, ce qui est louable.
Chapitre 13 : La pente descendante
L’épigénétique du vieillissement. Avec l’âge, l’épigénome se dégrade.
On observe une hypométhylation globale (le génome se "déverrouille" partout, permettant l’expression de gènes parasites ou de rétrovirus) et une hyperméthylation locale (certains gènes suppresseurs de tumeurs sont verrouillés par erreur, causant le cancer).
Horloge épigénétique de Horvath : il est possible de prédire l’âge d’une personne avec une précision de +/- 3 ans juste en regardant la méthylation de 353 sites CpG.
Vision très darwinienne : la sélection naturelle se moque de nous après 40 ans, donc les mécanismes de maintenance épigénétique ne sont pas parfaits. C’est une idée forte, bien expliquée.
Chapitre 14 : Longue vie à la reine
Étude de cas : l’abeille domestique.
La reine et les ouvrières sont génétiquement identiques. Pourtant, la reine vit 2 ans et pond, l’ouvrière vit 6 semaines et est stérile.
Différence clé : la gelée royale.
La gelée royale contient des composés (acides gras) qui inhibent une enzyme modifiant les histones (HDAC). Résultat : le génome de la larve est remodelé, elle devient reine.
Une simple manipulation chimique de l’épigénome transforme une ouvrière en reine.
C’est l’exemple ultime du pouvoir de l’environnement sur le génome. Très visuel et marquant.
Chapitre 15 : La révolution verte
L’épigénétique chez les plantes.
Chez les plantes, la transmission épigénétique est extrêmement stable (pas de reprogrammation comme chez les animaux).
Exemple de la linaire (toadflax) : une fleur génétiquement normale qui, à cause d’une méthylation excessive, devient symétrique au lieu d’asymétrique. Ce trait est héréditaire.
Intérêt agronomique : créer des variétés de plantes résistantes à la sécheresse sans modification génétique.
Critique :
Carey souligne que ce qui est une exception chez l’humain (transmission épigénétique) est la norme chez la plante. Elle ouvre la porte aux biotechnologies vertes.
Chapitre 16 : Les voies de l’avenir
Conclusion et perspectives médicales.
Biomarqueurs : détecter le cancer plus tôt en cherchant de l’ADN méthylé anormalement dans le sang.
Thérapies épigénétiques : les premiers médicaments (Inhibiteurs de HDAC, comme la Vorinostat, et inhibiteurs de DNMT, comme l’Azacitidine) sont déjà sur le marché pour les lymphomes et les leucémies.
Contrairement à la thérapie génique, on ne change pas la séquence. On "efface le marqueur au crayon" plutôt que de réécrire le livre.
Vision optimiste et prospective. Plus de 10 ans après la parution du livre, on peut constater que ces thérapies ont effectivement révolutionné le traitement de certains cancers (notamment les lymphomes), mais elles restent décevantes pour les tumeurs solides. Carey ne pouvait pas le prédire, mais son enthousiasme est mesuré.

"Handbook of Epigenetics: The New Molecular and Medical Genetics" (Trygve Tollefsbol, 2010) constitue le premier manuel de référence exhaustif qui va structurer la discipline elle-même. Un Ouvrage collectif dirigé, 37 chapitres, 624 à 639 pages selon les éditions, rédigés par des leaders internationaux du terrain, son plan est celui d'une discipline constituée : fondamentaux, méthodes, modèles, applications, pathologies, thérapies. Le communiqué officiel de l'Université d'Alabama à Birmingham (UAB), daté du 24 novembre 2010, est explicite. Tollefsbol lui-même déclare que cette publication est "the first to offer a fully comprehensive analysis of epigenetics". En 2010, aucun ouvrage n'avait tenté une couverture aussi exhaustive. Ce n'est pas un livre sur l'épigénétique, c'est le livre qui a institué l'épigénétique comme domaine autonome disposant de son propre manuel de référence. Le Handbook a fait l'objet d'une recension dans CHEMBIOCHEM (revue de la Chemistry Europe, facteur d'impact significatif). C'est donc un instantané historique. Indispensable pour comprendre comment le champ a été structuré conceptuellement, mais totalement insuffisant pour une pratique de recherche ou clinique actuelle...

Eva Jablonka & Marion J. Lamb, "Epigenetic Inheritance and Evolution: The Lamarckian Dimension" (Oxford University Press, 1995).
Eva Jablonka est une biologiste/théoricienne de l’évolution (Tel Aviv University), figure majeure des débats sur l’héritabilité épigénétique et, plus largement, sur une vision pluraliste de l’hérédité. Marion J. Lamb (biologiste, Birkbeck, University of London) a été la collaboratrice principale de cette dernière; elle est reconnue comme pionnière de l’épigénétique évolutive.
Epigenetic Inheritance and Evolution est un ouvrage précurseur: il remet au centre du débat, dès 1995, une question que la Synthèse moderne avait largement marginalisée : dans quelles conditions des variations induites par l’environnement / acquises peuvent-elles laisser une trace héréditaire, et quel pourrait être leur poids dans l’évolution. Il traite ainsi explicitement de « l’héritabilité des caractères acquis » et discute les arguments pour/contre, en s’appuyant sur des exemples et sur les connaissances de l’époque.
Le sous-titre "The Lamarckian Dimension" signale l’ambition : réexaminer (sans tomber dans le lamarckisme naïf) la possibilité d’une dimension “lamarckienne” via des mécanismes non strictement génétiques. L’ouvrage est fortement cité et a suscité des débats philosophiques et théoriques dès la fin des années 1990 (critiques et discussions dans Biology & Philosophy, par ex.). Il emploie alors le terme d' « épigénétique » au sens large (hérédité non-ADN, états cellulaires, etc.) : il n’est pas strictement aligné sur l’usage “marques de chromatine & séquençage” popularisé ensuite dans les années 2000. (C’est aussi ce qui a fait sa force à l’époque : rouvrir le dossier conceptuel.). Certains comptes rendus ont pointé des points discutables (sélection des exemples, débats sur “mutations dirigées” en bactéries, etc.), ce qui montre justement qu’il a été un détonateur de controverses plutôt qu’un consensus.

Eva Jablonka & Marion J. Lamb, "Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life" - (MIT Press, 2005 (hardcover) ; édition révisée et augmentée 2014).
Les idées sur l'hérédité et l'évolution connaissent actuellement une révolution. De nouvelles découvertes en biologie moléculaire remettent en question la version darwinienne centrée sur les gènes, selon laquelle l'adaptation ne se produit que par la sélection naturelle de variations aléatoires de l'ADN.
Dans "Evolution in Four Dimensions" , Eva Jablonka et Marion Lamb soutiennent que l'hérédité ne se limite pas aux gènes.
Elles identifient quatre « dimensions » de l'évolution : quatre systèmes d'hérédité qui jouent un rôle dans la transmission génétique, épigénétique (ou transmission cellulaire des traits non liés à l'ADN), comportementale et symbolique (transmission par le langage et d'autres formes de communication symbolique). Selon elles, ces systèmes peuvent tous fournir des variations sur lesquelles la sélection naturelle peut agir.
1️. - L’hérédité génétique
C’est le système classique de la biologie évolutive.
- support : séquence de l’ADN
- mécanismes : mutations, recombinaisons, ségrégation
- transmission : parents → descendants via les gamètes
- rôle évolutif : variations génétiques aléatoires soumises à la sélection naturelle
C’est la première dimension, centrale dans le néo-darwinisme traditionnel.
2️. - L’hérédité épigénétique
Elle concerne la transmission de traits sans modification de la séquence d’ADN.
- support : méthylation de l’ADN, modifications des histones, états de la chromatine
- mécanismes : maintien d’états d’expression génique au cours des divisions cellulaires
- transmission : cellules mères → cellules filles, parfois d’une génération à l’autre
- rôle évolutif : stabilisation de phénotypes induits par l’environnement
Elle explique comment des cellules ou organismes identiques génétiquement peuvent conserver des différences héritables.
3️. - L’hérédité comportementale
Elle repose sur la transmission sociale de comportements.
- support : apprentissage, imitation, traditions
- mécanismes : apprentissage social, parental, culturel
- transmission : individus → individus (au sein ou entre générations)
- rôle évolutif : diffusion rapide de comportements adaptatifs
Très importante chez les animaux sociaux (oiseaux, primates, mammifères marins).
4️. - L’hérédité symbolique
Spécifique à l’espèce humaine (ou quasi).
- support : symboles, principalement le langage
- mécanismes : enseignement, communication symbolique, institutions
- transmission : culturelle et cumulative
- rôle évolutif : transformation rapide et cumulative des comportements, techniques et organisations sociales
C’est la dimension qui permet une évolution culturelle cumulative, largement indépendante des changements biologiques lents.

"Evolution in Four Dimensions" offre ainsi une vision plus riche et plus complexe de l'évolution que la vision unidimensionnelle basée sur les gènes que beaucoup soutiennent aujourd'hui.
La nouvelle synthèse proposée par Jablonka et Lamb montre clairement que les changements induits et acquis jouent également un rôle dans l'évolution.
Après avoir discuté en détail chacun des quatre systèmes d'hérédité, Jablonka et Lamb « remettent Humpty Dumpty sur pied » en montrant comment tous ces systèmes interagissent.
Ils examinent comment chacun d'entre eux a pu voir le jour et guider l'histoire de l'évolution, et discutent des implications sociales et philosophiques de la vision quadridimensionnelle de l'évolution. ..
1️. - Implications philosophiques
- Remise en cause du réductionnisme génétique
L’une des implications majeures est le rejet du “tout-gène” : le gène n’est plus l’unique unité causale de l’évolution; les explications purement génétiques des traits complexes deviennent insuffisantes; l’évolution est un processus multi-niveaux, impliquant cellules, organismes, comportements et cultures. Ce qui conduit à une biologie plus systémique, où les causes sont distribuées et interactives.
- Redéfinition de l’hérédité
L’hérédité n’est plus uniquement verticale (parents → descendants), ni exclusivement matérielle (ADN), mais aussi cellulaire, sociale, culturelle; et parfois rapide, réversible et contextuelle. Philosophiquement, cela affaiblit la frontière stricte entre inné et acquis.
- Réhabilitation partielle de l’acquis
Sans revenir au lamarckisme naïf, Jablonka et Lamb montrent que certaines caractéristiques acquises peuvent être transmises; l’environnement peut induire des variations héréditaires; l’évolution n’est pas uniquement aveugle au sens fonctionnel.
L’évolution n’est plus seulement fondée sur le hasard + sélection, mais aussi sur des processus orientés par l’interaction avec l’environnement.
- Pluralité des mécanismes évolutifs
La sélection naturelle reste centrale, mais elle n’est plus le seul moteur explicatif : l’instruction, la plasticité, l’apprentissage et la symbolisation jouent un rôle causal. Ce qui enrichit le darwinisme plutôt que de l’abolir, en conduisant à un darwinisme élargi.
2️. - Implications pour la compréhension de l’humain
- Nouvelle lecture de l’évolution humaine : chez l’humain, l’hérédité symbolique (langage, normes, institutions) devient un facteur évolutif majeur; l’évolution biologique et l’évolution culturelle sont co-imbriquées. L’évolution humaine est en partie auto-influencée, via la culture.
- Fin du déterminisme biologique fort : puisque plusieurs systèmes interagissent, les comportements humains ne sont pas rigidement déterminés par les gènes; la plasticité, l’apprentissage et la culture modulent profondément les trajectoires individuelles. C'est s’opposer aux lectures biologisantes du comportement (sexisme, racisme, essentialisme).
3️. - Implications sociales et politiques
- Responsabilité collective accrue : si l’environnement social et culturel influence l’hérédité comportementale et symbolique, et peut même, indirectement, affecter des trajectoires biologiques, alors les sociétés ont une responsabilité évolutive vis-à-vis : de l’éducation, des conditions de vie et des structures sociales.
- Critique des usages idéologiques du darwinisme : la vision quadridimensionnelle fragilise les justifications biologiques des inégalités sociales, remet en question le darwinisme social implicite, et montre que la sélection n’est ni moralement normative ni socialement prescriptive. Ce n’est pas parce qu’un trait existe qu’il est “naturel” ou “inévitable”.
4️. Implications épistémologiques
- Changement de cadre explicatif
Les auteures plaident pour des explications historiques plutôt que purement mécanistes, une biologie attentive aux processus, pas seulement aux entités, et une interdisciplinarité forte (biologie, sciences sociales, philosophie). La biologie évolutive devient une science plus narrative, pluraliste et intégrative.
"(Prologue) ... Notre thèse principale est que la manière dont la biologie pense l’hérédité et l’évolution est en train de subir un changement révolutionnaire. Ce qui émerge est une nouvelle synthèse, qui remet en question la version néo-darwinienne centrée sur le gène, laquelle a dominé la pensée biologique au cours des cinquante dernières années.
Les changements conceptuels en cours reposent sur des connaissances issues de presque toutes les branches de la biologie, mais notre attention se portera ici principalement sur l’hérédité. Nous soutiendrons que :
- il y a plus dans l’hérédité que les seuls gènes ;
- certaines variations héréditaires n’ont pas une origine aléatoire ;
- certaines informations acquises sont héritées ;
le changement évolutif peut résulter de processus d’instruction autant que de sélection.
Ces affirmations peuvent paraître hérétiques à quiconque a appris la version classique de la théorie darwinienne de l’évolution, selon laquelle l’adaptation résulte de la sélection naturelle de variations génétiques dues au hasard. Néanmoins, elles reposent solidement à la fois sur de nouvelles données et sur de nouvelles idées. La biologie moléculaire a montré que nombre des anciennes hypothèses concernant le système génétique — fondement de la théorie néo-darwinienne actuelle — sont incorrectes. Elle a également révélé que les cellules peuvent transmettre des informations à leurs cellules filles par des mécanismes d’hérédité non fondés sur l’ADN (épigénétiques).
Cela signifie que tous les organismes possèdent au moins deux systèmes d’hérédité. De plus, de nombreux animaux transmettent des informations à d’autres individus par des moyens comportementaux, ce qui constitue un troisième système d’hérédité. Quant à nous, êtres humains, nous en possédons un quatrième, car l’hérédité symbolique — en particulier le langage — joue un rôle majeur dans notre évolution. Il est donc erroné de penser l’hérédité et l’évolution uniquement en termes de système génétique. L’hérédité épigénétique, comportementale et symbolique fournit également des variations sur lesquelles la sélection naturelle peut agir.
Lorsque l’on prend en compte les quatre systèmes d’hérédité ainsi que leurs interactions, une vision très différente de l’évolution darwinienne émerge. Cette perspective peut atténuer la frustration que ressentent de nombreuses personnes face à l’approche dominante centrée sur le gène, car il n’est plus nécessaire d’attribuer l’évolution adaptative de chaque structure ou activité biologique — y compris le comportement humain — à la seule sélection de variations génétiques aléatoires, aveugles à la fonction. Lorsque l’on considère tous les types de variations héréditaires, il devient clair que les changements induits et acquis jouent eux aussi un rôle dans l’évolution.
En adoptant une perspective à quatre dimensions, il devient possible d’élaborer une théorie de l’évolution bien plus riche et sophistiquée, dans laquelle le gène n’est plus l’unique cible de la sélection naturelle.
Nous avons divisé ce livre en trois parties, chacune précédée d’une brève introduction.
La partie I est consacrée à la première dimension de l’hérédité et de l’évolution : le système génétique. Dans le chapitre 1, nous retraçons l’histoire de la théorie de Darwin et montrons comment elle est devenue si fortement centrée sur le gène. Le chapitre 2 décrit comment la biologie moléculaire a transformé la manière dont les biologistes conçoivent la relation entre les gènes et les caractères. Dans le chapitre 3, nous examinons les données suggérant que toutes les modifications génétiques ne doivent pas être considérées comme des événements purement aléatoires.
La partie II traite des autres dimensions de l’hérédité. Le chapitre 4 porte sur la deuxième dimension, l’hérédité épigénétique, grâce à laquelle des cellules possédant un ADN identique sont capables de transmettre leurs caractéristiques à leurs cellules filles. Dans le chapitre 5, nous explorons les mécanismes par lesquels les animaux transmettent leurs comportements et leurs préférences par l’apprentissage social, constituant ainsi la troisième dimension. La quatrième dimension est abordée dans le chapitre 6, qui décrit la transmission de l’information par le langage et d’autres formes de communication symbolique.
Dans la partie III, nous reconstituons l’ensemble, à la manière de Humpty Dumpty. Après avoir examiné chacune des quatre dimensions de l’hérédité plus ou moins isolément, nous les réunissons en montrant comment, à long terme, les systèmes d’hérédité dépendent les uns des autres et interagissent (chapitres 7 et 8). Dans le chapitre 9, nous discutons de leur origine possible et de la manière dont ils ont orienté l’histoire évolutive. Enfin, dans le chapitre 10, nous résumons notre position et l’inscrivons dans une perspective plus large, en abordant certaines implications philosophiques de la vision à quatre dimensions, ainsi que des enjeux politiques et éthiques.
Chaque chapitre se termine par un « dialogue », et l’intégralité du chapitre 10 adopte cette forme. Nous utilisons ces dialogues comme un dispositif permettant de revenir sur certains points délicats de notre argumentation et de mettre en lumière les zones d’incertitude ainsi que les questions controversées. Les participants à ces dialogues sont M.E. (qui représente les auteures, Marion Lamb et Eva Jablonka) et un interlocuteur que l’on pourrait qualifier d’« avocat du diable », mais que, afin d’éviter les connotations négatives de ce terme, nous avons choisi d’appeler Ipcha Mistabra (I.M.). Ipcha Mistabra est une expression araméenne signifiant « l’hypothèse opposée » ..."
La vision quadridimensionnelle de Jablonka et Lamb enrichit profondément la théorie de l’évolution en mettant en évidence la pluralité des systèmes d’hérédité. Toutefois, si son apport est désormais reconnu pour l’épigénétique et le développement, son extension aux dimensions comportementale et symbolique demeure débattue, en raison de difficultés conceptuelles, empiriques et méthodologiques.
L'idée principale de Jablonka et Lamb n’est pas de rejeter Darwin, mais de pluraliser les supports d’hérédité et donc les sources de variation : à côté de l’hérédité génétique, elles insistent sur l’épigénétique, l’hérédité comportementale (apprentissage social) et l’hérédité symbolique (langage, systèmes de signes), toutes susceptibles de produire des différences transmissibles sur lesquelles la sélection peut agir.

Chez Ernst Mayr, la perspective est différente, car il demeure un architecte majeur de la Synthèse moderne. Dans "What Evolution Is" (Basic Books, 2001), Mayr met l’accent sur l’explication darwinienne « classique » : la variation apparaît dans les populations, puis la sélection (et d’autres processus) en modifie les fréquences, et l’essentiel du cadre explicatif reste populationnel et génétique. Son apport décisif — particulièrement dans ses ouvrages de synthèse historique comme "The Growth of Biological Thought: Diversity, Evolution, and Inheritance" (Harvard University Press/Belknap, 1982) — consiste à clarifier les grands concepts (pensée populationnelle, importance des isolats reproductifs, rôle de la contingence historique).
Face à Jablonka & Lamb, Mayr serait typiquement mobilisé pour rappeler que l’évolution, comme théorie scientifique, gagne en robustesse lorsque ses mécanismes sont définis de façon testable et quantifiable (variation, héritabilité, différentiel de reproduction). La critique « mayrienne » la plus naturelle envers la vision quadridimensionnelle n’est pas qu’elle est fausse, mais qu’elle risque de diluer l’hérédité (et donc l’explication évolutive) si l’on ne spécifie pas clairement quand et à quelles échelles les transmissions non génétiques sont suffisamment stables pour peser sur le long terme.

Richard Dawkins, lui, ne défend pas seulement la Synthèse moderne : il en propose une reformulation très influente, centrée sur le gène comme unité privilégiée d’explication. Dans "The Selfish Gene" (Oxford University Press, 1976 ; éd. anniversaires ultérieures), il popularise l’idée que la sélection naturelle se comprend puissamment en considérant les gènes comme des réplicateurs, les organismes étant des « véhicules » temporaires.
Et dans "The Extended Phenotype: The Long Reach of the Gene" (éd. originale 1982), il étend cette perspective en soutenant que les effets des gènes peuvent se manifester au-delà du corps (ex. constructions, modifications de l’environnement).
Comparé à Jablonka & Lamb, Dawkins est l’interlocuteur critique le plus direct ...
là où elles multiplient les systèmes d’hérédité, Dawkins tend à ramener beaucoup de phénomènes (y compris comportementaux) à des stratégies façonnées par la sélection de gènes, ou à des prolongements de leurs effets.
En version forte, l’objection « dawkinsienne » serait : les transmissions épigénétiques, comportementales ou symboliques peuvent exister, mais si elles n’ont pas une stabilité et une fidélité suffisantes, ou si elles ne s’ancrent pas in fine dans des différences génétiques affectant la reproduction, elles restent secondaires par rapport au moteur génétique de l’évolution.
La réplique de Jablonka & Lamb serait précisément que cette hiérarchisation est une hypothèse — pas une nécessité — et que, dans certains contextes, les systèmes non génétiques peuvent initier des trajectoires adaptatives.

Massimo Pigliucci, enfin, occupe une position intermédiaire qui permet souvent de « traduire » Jablonka & Lamb dans un langage accepté plus largement par l’évolution contemporaine.
D’un côté, Pigliucci a beaucoup travaillé sur la plasticité phénotypique ("Phenotypic Plasticity: Beyond Nature and Nurture", Johns Hopkins University Press, 2001), qui sape déjà l’opposition simple gènes/environnement et rend plausible l’idée de variations induites et stabilisées.
De l’autre, il est coéditeur avec Gerd B. Müller de Evolution—"The Extended Synthesis" (MIT Press, 2010), ouvrage emblématique de la Synthèse évolutionnaire étendue (EES), qui cherche à compléter la Synthèse moderne en intégrant davantage la biologie du développement, les biais développementaux, la plasticité, la construction de niche, etc. Dans ce cadre, Jablonka & Lamb apparaissent moins comme des « hérétiques » que comme des figures d’un même mouvement pluraliste : elles poussent très loin le pluralisme des héritages (jusqu’au symbolique), tandis que Pigliucci/Müller cherchent à formaliser une extension compatible avec les standards méthodologiques de la biologie évolutive.
La différence critique est alors surtout une différence de degré et de style théorique.
- Jablonka & Lamb assument une thèse forte ..
il existe plusieurs systèmes d’hérédité relativement autonomes, dont certains peuvent transmettre de l’information acquise, et l’évolution doit se penser « en quatre dimensions ».
- Pigliucci, même lorsqu’il est ouvert aux extensions, se montre souvent plus prudent sur ce qui peut être stabilisé, quantifié et intégré dans des modèles ; son projet vise à enrichir la théorie sans perdre en testabilité.
Autrement dit, Pigliucci est fréquemment lu comme celui qui « institutionnalise » une partie des intuitions de Jablonka & Lamb dans un programme de recherche plus consensuel (EES), tandis que Dawkins représente le pôle « gène d’abord », et Mayr le pôle « synthèse classique & clarification conceptuelle ».

L'épigénétique est un champ extraordinairement solide sur les mécanismes locaux (comment une cellule se souvient qu'elle est une cellule de peau). Elle est extraordinairement prudente et débattue sur les promesses de transmission héréditaire des traumatismes ou des comportements.
C'est précisément dans cette zone d'incertitude (l'hérédité) que se glissent les "mysticismes" que l'on traque ..
Ce qui est solidement établi
- La différenciation cellulaire est épigénétique : Une cellule du foie et un neurone ont le même ADN. Leurs différences sont maintenues par des marques épigénétiques stables .
- La réversibilité : Contrairement aux mutations, les marques épigénétiques peuvent être effacées (ex: reprogrammation des cellules souches) .
- L'impact environnemental : La nutrition, le tabac, le stress modifient effectivement les marques épigénétiques au cours de la vie d'un individu .
- L'application médicale : Des "épi-médicaments" sont déjà utilisés (notamment dans les leucémies). La méthylation est un biomarqueur pronostique validé pour certains cancers .
Les incertitudes et controverses majeures (Le "Front de science")
A. La transmission héréditaire (Le débat principal)
Chez les plantes : C'est prouvé et accepté .
Chez les mammifères (dont l'Homme) : C'est la zone d'incertitude maximale. Il existe une "remise à zéro" massive des marques épigénétiques lors de la formation des gamètes et de l'embryon .
Le consensus actuel : Si transmission il y a, elle est exceptionnelle, partielle, et son mécanisme exact (comment la marque "échappe"-t-elle à l'effacement ?) est débattu. On ne peut pas affirmer que les traumatismes de vos grands-parents sont inscrits "dans vos gènes" de manière certaine .
B. "L'héritable" n'est pas "l'hérité" (Erreur logique majeure)
Une source académique de bioéthique met en garde contre une confusion grave : en biologie moléculaire, "héritable" signifie simplement que la marque se transmet lors de la division d'une cellule (d'une cellule mère à ses deux cellules filles). Cela ne signifie pas qu'elle est "héritée" de votre grand-parent (transmission sexuée). Utiliser ces termes comme synonymes est une erreur scientifique fondamentale .
C. La part du déterminisme
L'épigénétique ne remplace pas la génétique. Elle la complète. Parler d'une "révolution" qui annulerait Darwin ou le "tout génétique" est un discours radical non représentatif de la recherche actuelle .

L’épigénétique connaît des niveaux d’acceptation et des modes d’appropriation radicalement différents selon les pays.
Les résultats de recherche permettent d’identifier non pas un mais trois régimes distincts de « vogue » épigénétique, selon qu’elle est adoptée comme marché économique, comme priorité scientifique d’État, ou comme récit d’émancipation identitaire.
1. La « vogue » technico-économique : États-Unis, Allemagne, Chine
Dans ces pays, l’épigénétique est acceptée comme un secteur d’investissement prioritaire et un levier de compétitivité industrielle : États-Unis (Marché mondial n°1), Allemagne (Leader européen), Chine (croissance la plus rapide). L’épigénétique est acceptée comme technologie, pas comme philosophie. La « vogue » est portée par des marchés du diagnostic et du médicament (oncologie, hépatite, cardiologie). Le discours est celui de la performance thérapeutique et du retour sur investissement.
2. La « vogue » institutionnelle et sociale : France, Royaume-Uni, Europe du Nord
Dans ces pays, l’épigénétique est acceptée comme un outil de santé publique et de prévention environnementale. La « vogue » y est épistémique et politique.
- Royaume-Uni : 22% du marché européen. Le moteur est gouvernemental : UK Epigenetics Network, 1,5 Md£ alloués à la recherche génomique (2023) . L’acceptation y est intégrée à la stratégie de médecine de précision.
- France : 18% du marché européen . Mais le phénomène est plus profond qu’un simple chiffre. La France structure une école scientifique spécifique.
La France et l’Europe du Nord acceptent l’épigénétique car elle donne une base moléculaire aux inégalités sociales de santé. Elle permet de lier exposome, précarité et maladies chroniques. C’est une science qui résonne avec les valeurs de solidarité et de santé publique. L’acceptation y est institutionnalisée et politisée (au sens noble).
3. La « vogue » épistémique et identitaire : Australie autochtone
C’est le cas le plus singulier, et le plus révélateur de votre question sur « l’acceptation différentielle ».
- Australie – Communautés autochtones :
Un projet de recherche de l’Université Deakin (2023) étudie pourquoi les peuples autochtones australiens adoptent massivement et positivement le discours épigénétique .
Raisons profondes de cette « vogue » spécifique : l’idée que les traumatismes historiques s’inscrivent dans le corps et se transmettent résonne avec les récits autochtones de guérison intergénérationnelle. L’épigénétique est perçue comme une science anti-eugéniste, qui « dissout les frontières entre social et biologique » et peut légitimer des revendications de réparation. Cette acceptation enthousiaste comporte un risque de « re-biologisation de la race » , où la souffrance historique serait essentialisée dans l’ADN .
L’épigénétique n’est pas acceptée malgré son incertitude scientifique (transmission héréditaire), mais à cause d’elle. La zone d’ombre devient une ressource narrative et politique.
L’épigénétique n’est pas acceptée de la même façon partout
car elle ne répond pas aux mêmes questions selon les pays ...
- Aux États-Unis, elle répond à la question : Comment guérir le cancer ?
- En France, elle répond à : Comment prouver que la pollution rend malade ?
- En Australie autochtone, elle répond à : Comment faire reconnaître scientifiquement un traumatisme colonial ?